
Light Cycle Controlled Feeding and Weight Changes in Pigeons
William L. Palya
Department of Psychology, Jacksonville State University, Jacksonville, AL 36265, USA
Philip Hurst
Department of Psychology, Auburn University, Auburn, AL 36830, USA
Joseph J. Kenny
Department of Psychology, Earlham College, Richmond, IN47374, USA
William M. Gardner and Gary Seale
Department of Psychology, Jacksonville State University, Jacksonville, AL 36265, USA
PALYA, W. L., P. HURST, J. J. KENNY, W. M. GARDNER and G. SEALE. 1983. Light cycle controlled feeding and weight changes in pigeons. Bird Behaviour 4:71-77.
Systematic changes in body weight and key-pecking for food were obtained in pigeons as a result of light cycle manipulations. The first experiment demonstrated that there was an increase in body weight and in the rate of an operant which produced food just before the dark period in a day with 16 h of light and 8 h of darkness. It also demonstrated that some birds exhibited long term fluctuations in body weight of about 8% to 9%. In the second experiment pigeons were maintained under five different circadian light cycles. An increase in feeding just before the dark period was obtained in each light cycle. Constant illumination abolished the characteristic feeding pattern in both experiments, and the feeding pattern was substantially altered in both when feeding during a portion of the 24 h cycle was precluded by removing food rather than by removing light. Both experiments also demonstrated that when maintained under constant illumination the birds earned fewer reinforcements but paradoxically weighed more than when maintained under a light cycle.
Light cycles Feeding Body weight Pigeons
Introduction
Feeding strategies are an important aspect of adaptive behaviour. Therefore, knowledge of the factors that govern feeding should help us to understand behaviour in general and even to understand the rules for assuring generality between animal research and human behavior.
The context in which food is available can have an important influence on feeding behaviour. Most notable among these effects is the modulation of feeding by rhythmic factors such as light. Siegel (4) observed the effect of a natural circadian light cycle on feeding by rats. He found that food intake was lowest in the light period; that it was highest early in the dark period; and that it exhibited a secondary peak later in the dark period. Zeigler, Green and Lehrer (5) observed a similar circadian feeding pattern in pigeons. They found that with unlimited food feeding occurred in bouts, and that feeding occurred early in the light period and again late in the light period. The birds rarely ate in total darkness.
The present study determines whether light changes are responsible for the circadian pattern in feeding by pigeons. This would verify the hypotheses of Murton (2) and Zeigler et al., (S) who suggested that the pattern of feeding was controlled by the light cycle. We wished to determine whether periodic access to food under constant illumination would create similar feeding patterns, and whether equivalent feeding patterns could be maintained under various light cycles but abolished under constant illumination. Cycles of more or of less than 24 h could not be adequately controlled and were therefore not used.
Experiment 1
This experiment investigated the effects of a typical 24 h light cycle on the feeding behaviour of pigeons. Feeding was measured by changes in body weight and for some birds by key-pecking that produced their only access to food. The experiment consisted of five sequentially administered treatments. Each experimental treatment or 'phase' was continued until the dependent measure appeared stable. This assured that calculations, such as a mean taken over the last five days of a phase, were always based on stable data.
Methods
This experiment used 20 Birmingham Rollers and four Modena pigeons. They were of unknown sex and between eight months and two years old, but without experimental histories. The pigeons were housed in commercial open-wire cages of 27 x 27 x 24 cm deep that were located in a 12 square metre windowless colony room. The animal room was closed to general access during the dark cycle, and weighings were done in a separate room.
All birds were initially exposed to an L:D 18:6 light cycle (Phase 1). This was followed by a period of constant illumination (Phase 2) to determine whether the light cycle was responsible for the pattern of feeding obtained in Phase 1. Food was then periodically unavailable (Phase 3) to determine if the actual controlling factor in Phase 1 was the absence of food when birds were in darkness rather than the light cycle. During this phase, food was unavailable during the period that was comparable to the dark portion of the L:D 16:8 light cycle (F:F 16:8). All pigeons were then returned to constant illumination (Phase 4) and finally to the L:D 18:6 light cycle (Phase 5) to assure that the obtained findings were not simply some cumulative result of the procedure, such as continual exposure to being fed in cages. Table I presents the various light cycles used and the number of days of each.
Sixteen pigeons were used to assess the effects of light cycle on body weight. The pigeons had free access to food, water and grit.
TABLE I
SUMMARY OF PHASES IN EXPERIMENT 1.
Phase (L:D) Days Per Phase
1 16:8 12
2 24:0 27
3 24:0 (F:F 16:8) 19
4 24:0 9
5 16:8 10
They were weighed every 3 h. Four of the pigeons were used to assess the effects of light cycle on feeding as measured by key-pecking that produced their only access to food. These birds were required to peck 10 times to produce a presentation of food for 4 s (called a chronic FR10 schedule of reinforcement). The cages containing these four pigeons were modified to accommodate aluminum response panels measuring 30 cm wide by 33 cm high. A 5 cm diameter food aperture was medially located on the response panel 9 cm above the wire floor. A 2 cm diameter key which required 0.15 N (Newton) force to operate was located 9 cm above the food aperture and 9 cm to the right. Each key was dimly lit white at all times except during the phase of restricted food. Water and grit were freely available and were provided in cups adjacent to the response panel.
Results
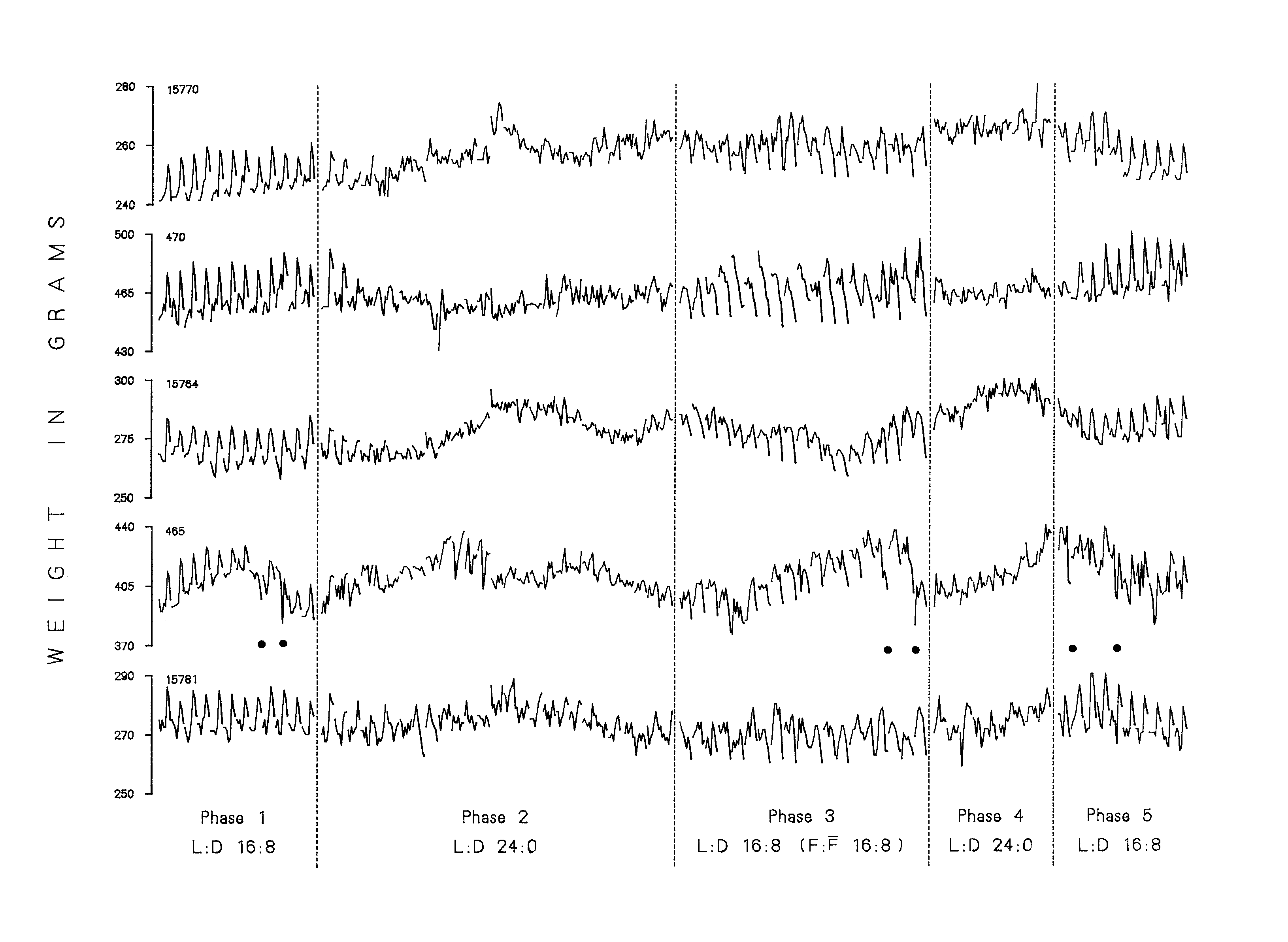
Figure 1. Daily weights for five sample birds based on recordings every 3 h for 77 days. The eight weights from each 24 h period are connected. Dots below the tracing for Bird 465 represent eggs.
Figure I presents the body weights obtained every three hours for 77 days for five sample birds. Records for each of the 16 birds were similar with respect to the weight changes which occurred within each day. However, not all birds maintained their average daily weight within as narrow range as might be expected. As can be seen in Figure 1, long-term changes in body weight of 8%or 9%occurred in some cases. The records were selected to illustrate the types of long term changes that were obtained. The weight record for bird 465 is included to illustrate the weight changes associated with egg laying. The filled circles below this record indicate the occurrence of eggs. Small breaks appear in the weight records of each bird between different 24 h cycle while dotted lines separate experimental treatments. A break in the middle of Phase 2 indicates a 14 day period when no weights were obtained.
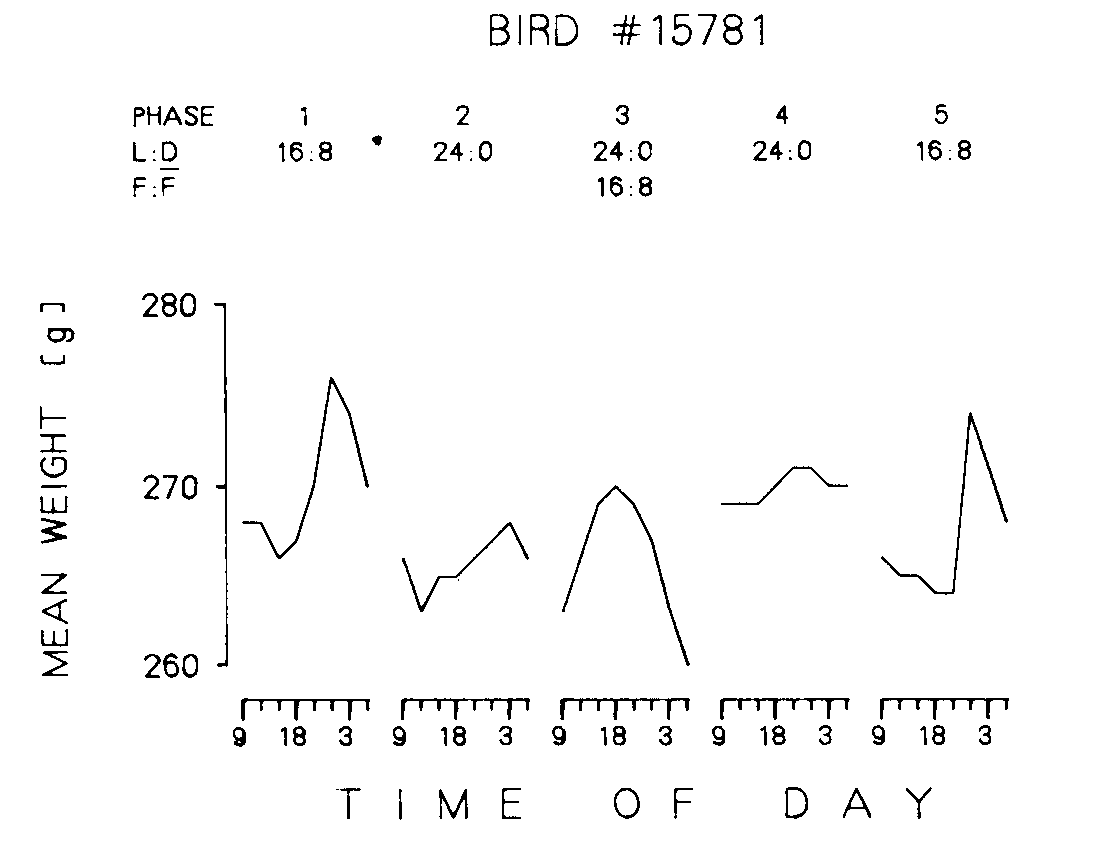
Figure 2. Mean daily weights for bird 15781 for 3 h intervals over the last five days of each phase.
Figure 2 presents the mean weight of a representative bird (15781) at each 3 h weighing throughout the day for each of the five phases. As can be seen the average body weight sharply increased immediately before the dark portion of an L:D 16:8 (Phase I and 5); whereas the cycle with 16 h of food and 8 h of no food (Phase 3) resulted in an increase followed by a decrease in body weight. Under continuous light (Phases 2 and 4), weight fluctuated much less throughout the day. By comparing Figure 2 with the line segments for this bird in Figure 1, it can be seen that these averaged data do not misrepresent the raw data.
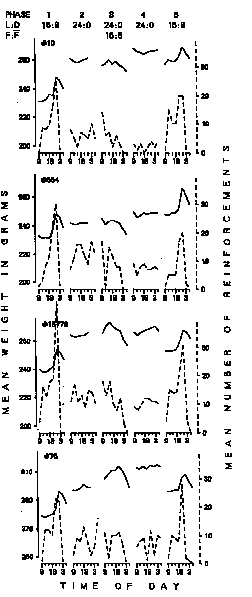
Figure 3. Mean daily weights (solid lines) and mean daily number of reinforcements (dotted lines) for four birds in response contingent group at 3 h intervals over last five days of each phase.
Figure 3 displays the data obtained from the birds which were required to key-peck for food: the average weight and average number of reinforcements for each 3 h period for each phase are presented. The figure shows that the daily distribution of obtained reinforcements roughly paralleled the weight changes. There was typically a sharp increase in weight and number of reinforcements immediately before the dark portion of the cycle (Phases I and 5). When the birds were exposed to constant illumination (Phases 2 and 4), their weight increased only slightly or remained relatively constant, and the distribution of obtained reinforcements similarly showed no consistent trend. Removal of food for eight of the 24 h in Phase 3 affected the group of birds that had to work for food in about the same way as the group of birds that were normally maintained with free access to food. Body weights tended to increase then decrease, or remain stable throughout the day, while the number of reinforcements obtained in each 3 h block tended to decrease or remain stable across the day. It can be seen by comparing Figure 2 and Figure 3 that a broadly similar pattern of weight change occurred in both the birds on a chronic FR schedule and in free feeding birds.
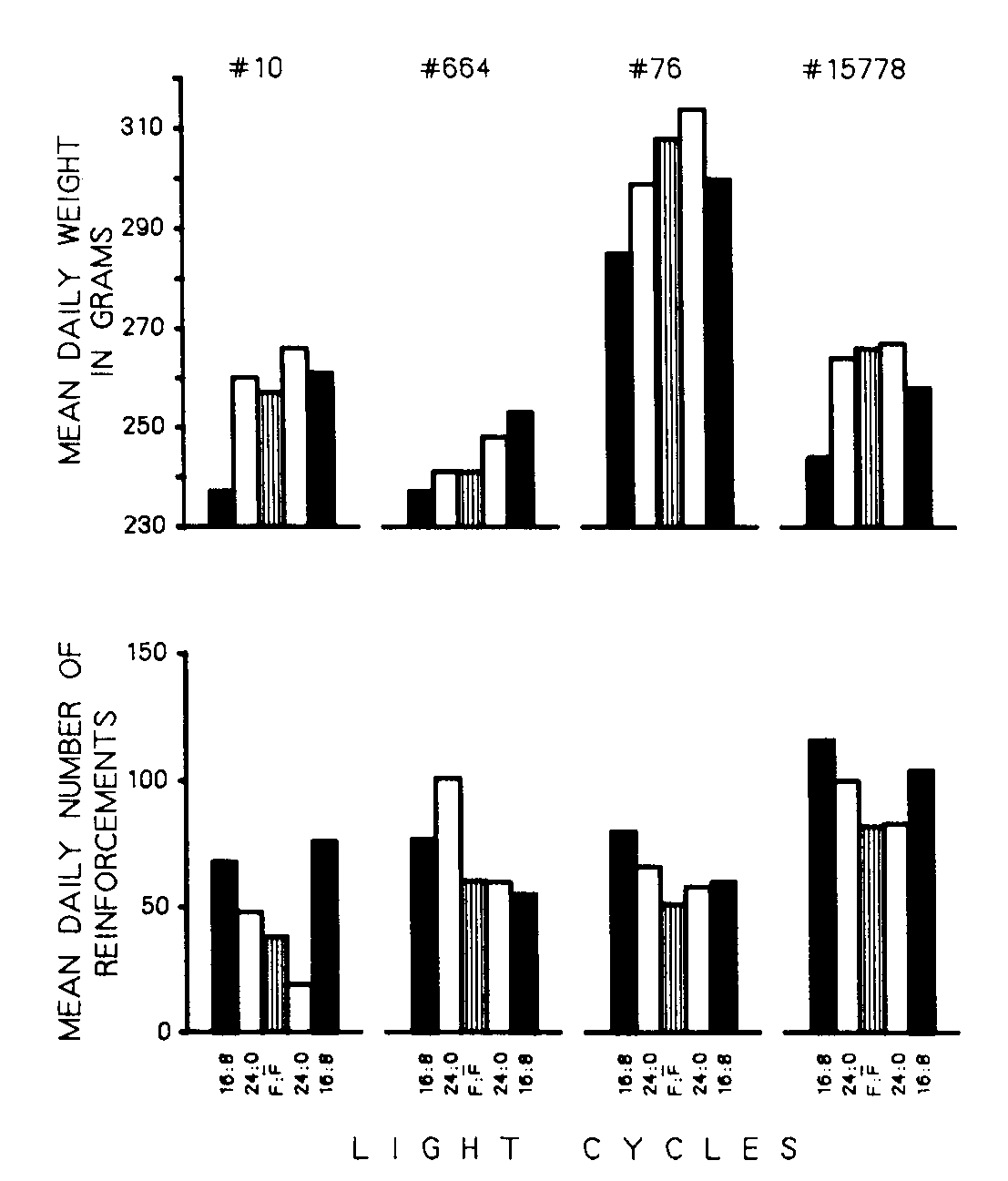
Figure 4. Mean weight and mean number of reinforcements over last five days of each phase for each bird in response contingent group.
In addition to affecting the distribution of feeding during the day, the light cycle also affected other aspects of feeding. The pigeons weighed more when maintained under continuous illumination but, paradoxically, produced fewer reinforcements per day. Figure 4 demonstrates this by presenting the mean daily weight and mean number of reinforcements for each bird over the last five days of each phase. As can be seen in the four sets of the upper part of Figure 4, almost every change between a light cycle and continuous illumination indicates that weights were greater under continuous illumination. However, the lower position indicates that three out of the four birds obtained more food under a light cycle than under continuous illumination.
Experiment 2
The first experiment assessed the effect that an L:D 16:8 light cycle had on feeding as indicated by daily changes in body weight and number of reinforcements. Experiment 2 also examined the effects of light cycle on feeding as indicated by the distribution of obtained reinforcements throughout the day. In this case, however, the pattern of feeding that occurred under several different 24 h light cycles was assessed. Each light cycle was imposed until stability was obtained on the keypecking measure. Before the birds were exposed to a different light cycle, they were maintained under continuous illumination until stability was again obtained on the key-pecking measure.
Methods
In this experiment we used four additional adult pigeons of unknown sex. All were housed in a battery of four contiguous open-wire cages in a light-tight room throughout the experiment. Each cage was 34 cm by 40 cm by 44 cm deep. The fronts were modified to accommodate aluminum response panels measuring 34 cm high by 30 cm wide. Water cups immediately beside the response panels were kept full. A 5 cm diameter food aperture was medially located on the response panel 9 cm above the wire floor. A 2 cm diameter key which required 0.15 N force to operate was located 9 cm above the food aperture and 9 cm to the right. Each key was dimly lit white at all times except during Phase 14. General illumination was provided by a 60 W bulb in front of the cages and a 40 W fluorescent tube behind the cages.
All birds had uninterrupted access to the functioning response panels, the only source of food. Throughout the experiment, 50 key pecks were required to produce 4 s access to Purina Pigeon Pellets (a chronic FR5O schedule). Phase 14 did not manipulate the light cycle. It was implemented to assure that the distinctive pattern of responding obtained in the previous phases was the result of the light cycle itself rather than inaccessibility of food in the dark period. Consequently, during Phase 14 general illumination was provided continuously; however, the response panel was operative for only nine of the 24 h. The key light was dark when the panel was inoperative. Table 2 presents the light cycles associated with each phase of this experiment and the number of days each was in effect.
Results
The results of Experiment 2 paralleled the findings of Experiment 1. Light cycles caused a peak in eating which occurred just before the dark period, while continuous illumination control conditions resulted in a constant rate of eating throughout the day.
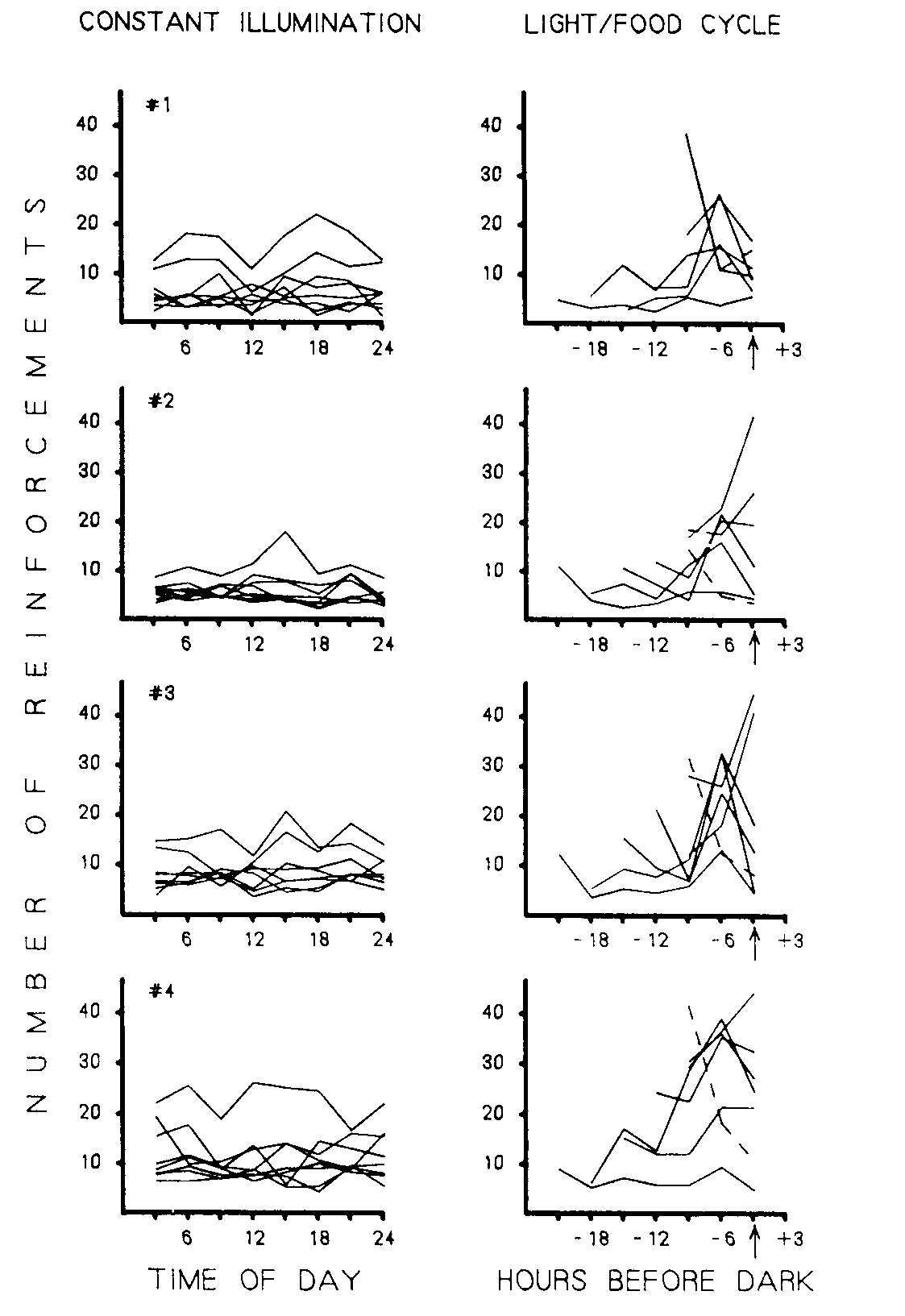
Figure 5. Number of food presentations earned by bird in each 3 h blocks for different light cycles. Mean daily number of reinforcements for 3 h intervals over the last five days of each phase is presented.
The daily distribution of obtained reinforcements under each of the seven experimental treatments and eight control conditions for each bird is presented in Figure 5. The data for each bird are presented in a horizontal pair of frames. The right frame presents data from light cycles in Phases 2, 4, 6, 8, 10 and 12 and the food cycle in Phase 14. While the left frame shows the distribution of food presentations during the intervening constant illumination control conditions (L:D 24:0). These were Phases 1, 3, 5, 7, 9, 11, 13 and 15. In each frame the abscissa represents consecutive 3 h periods, while the ordinate indicates the mean number of food presentations. Each of the 15 functions, therefore, indicates the daily feeding pattern which resulted from the conditions of one of the 15 phases. As can be seen in the left frame, constant illumination resulted in relatively evenly distributed feeding throughout the 24 h period in nearly all cases.
The light and food cycle frames present data in terms of hours before the lights were turned off or food was removed; therefore, a point on the abscissa does not necessarily indicate the same absolute time for each of the functions. The arrows beneath each frame indicate when the lights went out (or when food was removed in the case of the broken lines). A similar pattern of feeding across a wide range of light cycles with a 24 h base is evident in these frames. Typically more food presentations were obtained in the next to the last 3 h interval. However, more food was occasionally obtained in the final three hour period.
The result of periodic inaccessibility of food under continuous illumination was different. These functions (the broken lines) show a steady decrease in feeding throughout the day, rather than an increase followed by a decrease. The functions are similar from one bird to the next and are consistent with data from the similar manipulation in Experiment I .
A single exception to this summary occurred. Bird I under an L:D 9:15 (Phase 12) generated a function which was very similar to its food removal distribution of Phase 14.
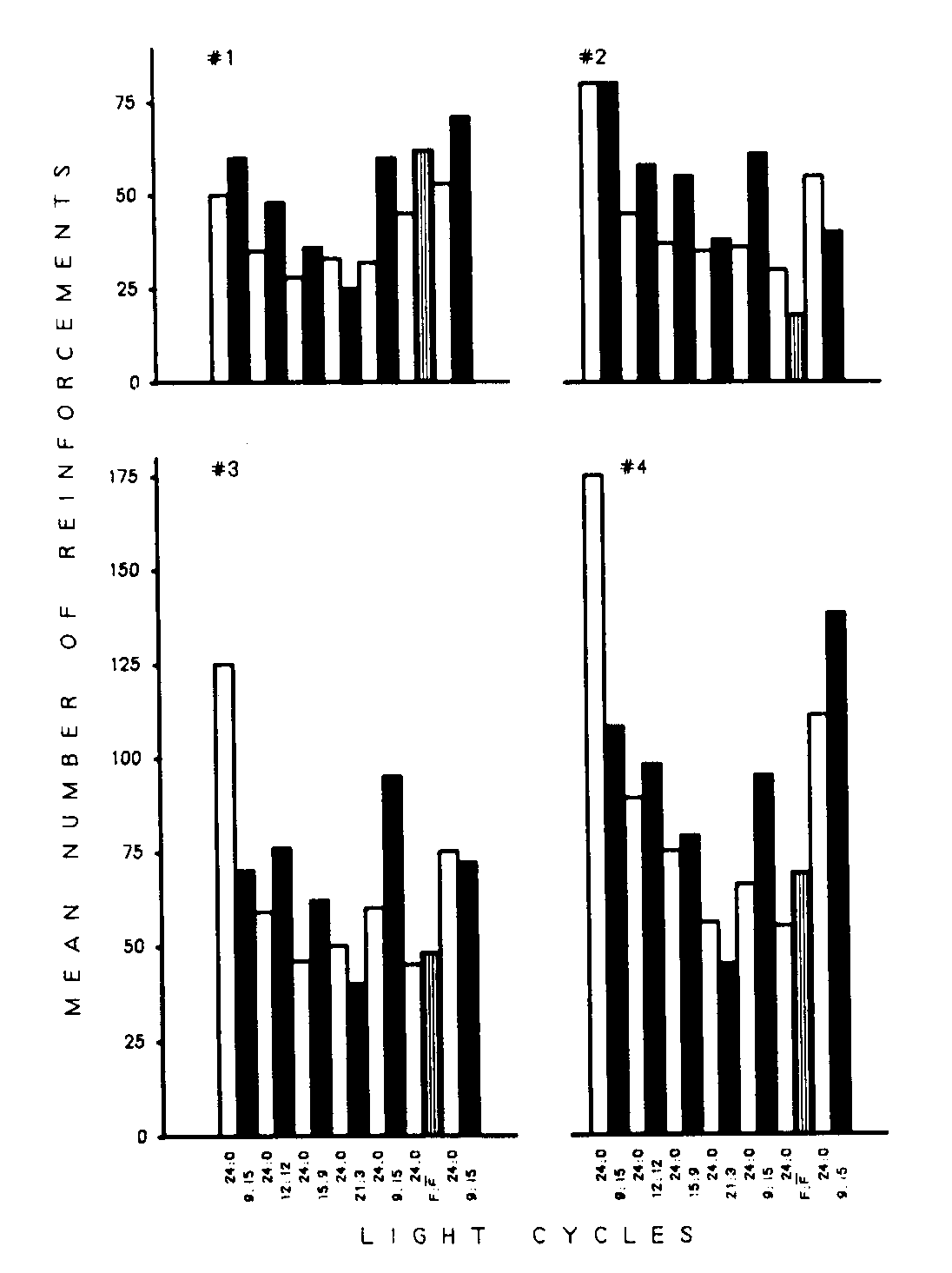
Figure 6. Mean number of reinforcements per day over the last five days of each phase for each bird.
The birds in this experiment typically obtained fewer reinforcements under constant illumination than when under a light cycle. This extends the similar findings in Experiment I to a variety of light cycles. The four frames of Figure 6 present the mean number of reinforcements obtained per day during each of the 15 phases by each bird. The exceptions to the general rule, that birds obtain more reinforcements when under a light cycle, are most obvious in Phase I and in the L:D 21:3 light cycle.
Discussion
Pigeons exhibit a sharp increase in feeding at the end of the illuminated portion of a light cycle. This pattern of feeding could be seen in both weight changes, and patterns of response on a chronic FR schedule. The distinctive pattern was also obtained under several circadian light cycles. The present study explicitly demonstrated that the pattern of feeding was controlled by the light cycle. It did this by documenting a characteristic pattern of feeding under a wide variety of circadian light cycles; by showing that the feeding pattern was abolished with constant light; and, by demonstrating that the feeding pattern could not be established by periodic removal of food under constant light. This study, therefore, extends the findings of Murton (2), Zeigler et al. (5) and Palya (3) who observed a peak in feeding just before darkness and suggested that light was responsible for that effect.
While it could be suggested that the weight changes actually reflect patterns of drinking or excretion, the distribution of earned reinforcements is not subject to that interpretation and supports the contention that feeding was in fact being monitored. It was unlikely that the observed feeding distribution was due to factors other than light. Similar patterns occurred with light cycles as short as 9:15 and as long as 21:3, even though other cyclic factors, such as temperature or lab activity, were not correlated with specific portions of the various light cycles. The control phases provide additional support for rejecting factors other than light. The feeding pattern was abolished under constant illumination and was nearly reversed by removing food, even though the other, potentially confounding, cyclic factors in the laboratory were not altered.
The observed patterns could not have been simply the result of truncating behaviour that occurred under constant light. The birds typically ate at a lower and constant rate under constant illumination and their feeding distributions over the early part of the day were not at all similar to those occurring under a light cycle. Neither was the feeding pattern a result of the simple absence of food. The consequences of having not eaten were identical in the light/no light and in the food/no food phases; yet the feeding distributions were markedly different. The light cycle itself, therefore, controlled a sharp increase in feeding just before dark. It is interesting to note that the increase in feeding followed light onset by an appropriate delay, even though the feeding rate increased only after many hours and that the peak in latency to feed was determined by the relative duration of light rather than simply a fixed delay. Although a homeostatic depletion-repletion model of motivated behaviour is the most parsimonious and popular explanation of feeding behaviour, it does not predict our data. This classical view suggests that feeding occurs when energy stores reach a critically low threshold, continues until some upper threshold is encountered and is then followed by fasting until the low threshold is again encountered. Collier, Hirsch and Kanarek (l) have argued for an alternative view of feeding behaviour. They point out that a simple threshold model is not consistent with findings such as the poor correlation between meal size and the pre-meal interval or the relatively small weight loss and maintenance of intestinal load across the intermeal interval. They also point out that reductionistic explanations that appeal to an inner process are as inappropriate for feeding behaviour as they were for operant behaviour. In both cases the appropriate explanation must be in terms of functional relationships between environmental manipulations and behaviour.
Acknowledgments
The authors gratefully acknowledge the contributions of Elizabeth Palya to all aspects of this research. Requests for reprints should be sent to William L. Palya.
1. Collier, G., E. Hirsch and R. Kanarek. 1977. The operant revisited. In: Handbook of Operant Behavior. Edited by W. K. Honig and J. E. R. Staddon. Englewood Cliffs, N.J.: Prentice-Hall.
2. Murton, R. K. 1965. The Wood Pigeon. London: Collins.
3. Palya, W. L. 1976. Light cycle controlled weight changes in pigeons. Journal of Interdisciplinary Cycle Research 7: 1 13-1 17.
4. Siegel, P. S. 1961. Food intake in the rat in relationship to the dark-light cycle. Journal of Comparative and Physiological Psychology 54:294-30 1.
5. Zeigler, H. P., H .L. Green and R. Lehrer. 1971. Patterns of feeding behavior in the pigeon. Journal of Comparative and Physiological Psychology 76:468-477.